|
|
 |
|
RECHERCHE Axes généraux Je suis chercheur systématicien : mon métier est, en sciences naturelles, de rendre compte de la diversité du vivant par sa structure et son histoire à travers les classifications phylogénétiques. Sous l’impulsion de Roland Billard et André Adoutte, dès 1989 j’ai commencé à construire ce qui deviendra à partir de fin 1993 avec Simon Tillier, Annie Tillier et Eric Pasquet le service de systématique moléculaire du MNHN, que j’ai animé dix années durant. Recruté comme assistant en 1988 dans un laboratoire d’Ichtyologie, j’ai travaillé en parallèle sur les téléostéens (les « poissons » modernes) et me suis même concentré sur la phylogénie de l’un des groupes de prédilection de membres de mon laboratoire d’alors, les notothénioïdes (téléostéens antarctiques). Les données que j’acquiers, avec mon équipe, sont majoritairement des données moléculaires ; mais les données de séquences ne m’intéressent vraiment que comme témoins de l’apparentement des espèces. Le reste de mon activité de recherche est tourné, pour une part minoritaire, vers la collecte des organismes sur le terrain (cinq missions effectuées en Antarctique) et, pour une part majoritaire, vers des questions méthodologiques et épistémologiques de la systématique. Distribution des thèmes de recherches par publications (les chiffres réfèrent à ceux de la page "publications profesionnelles") I. Méthodologie et exportation de l’analyse phylogénétique : 50% Ia.Problèmes
méthodologiques posés par la non congruence des phylogénies. Publications n°2, 3, 6,
28, 30, 46, 50, 58, 78, 84. II. Phylogénie des téléostéens : 50% IIa. Phylogénie
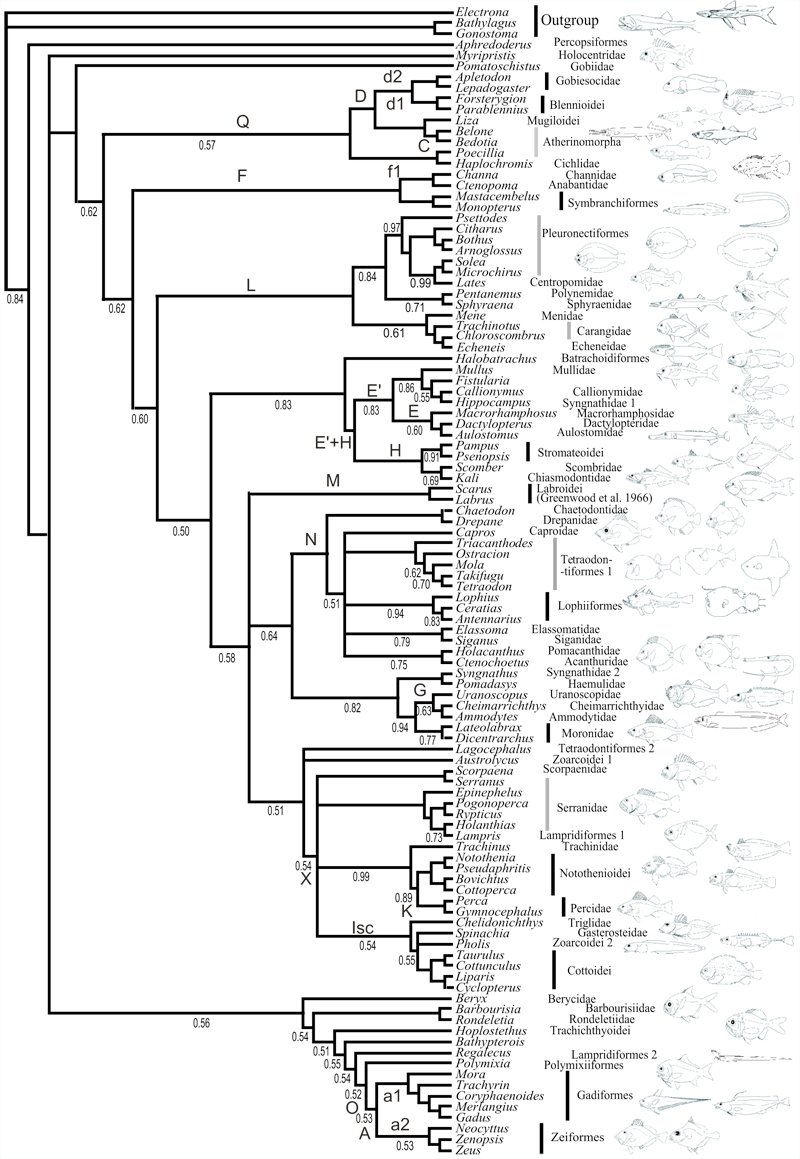
des Acanthomorpha (les acanthomorphes représentent 60% des téléostéens,
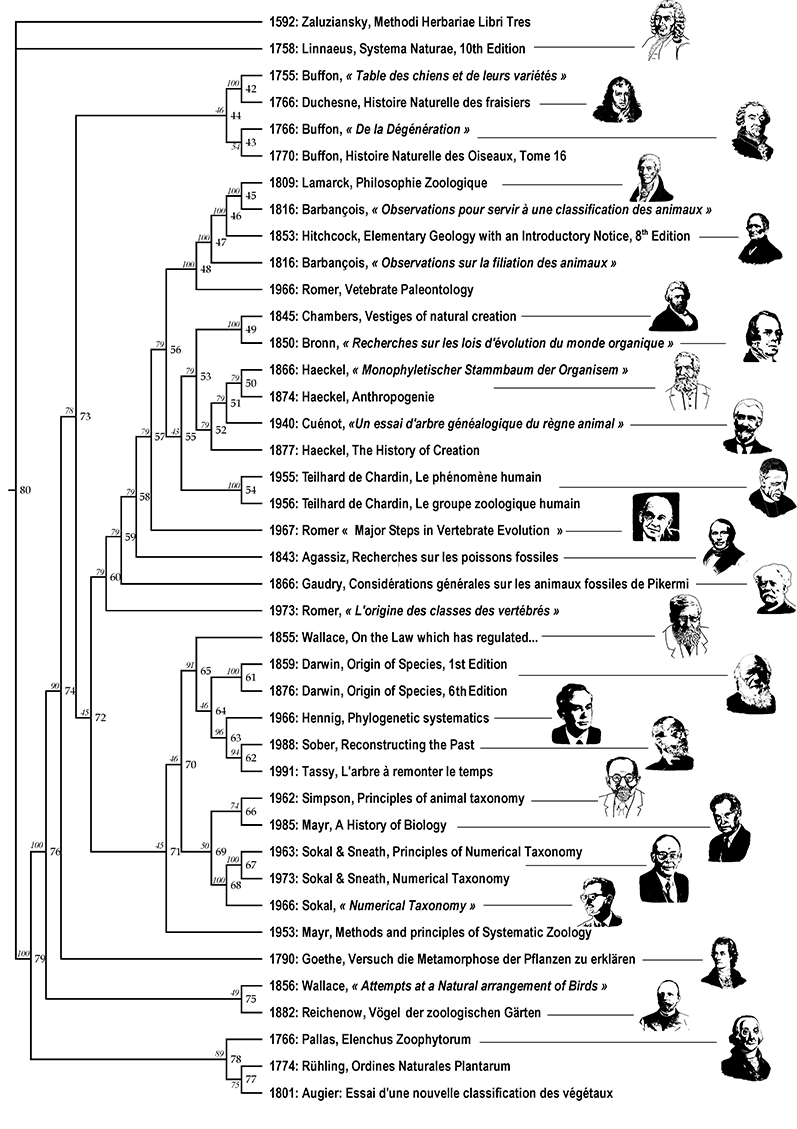
33% des vertébrés) : Publications n°52, 54, 59, 75, 77, 99, 100, 101. Des méthodes Mes méthodes sont celles de la phylogénie : méthodes standard de phylogénie moléculaire et phylogénie fondée sur des caractères anatomiques. Les questions de méthodes suivent deux axes. Le premier consiste à explorer comment construire des arbres phylogénétiques sans se tromper. En d’autres termes, qu’est-ce qui constitue une inférence phylogénétique fiable ? Presque tous mes articles de phylogénie traitent de cette question, d’une manière ou d’une autre, en plus de leurs conclusions zoologiques et/ou classificatoires. Avec Pierre Deleporte nous avons organisé en octobre 2004 le colloque du vingtième anniversaire de la société Française de Systématique autour du thème « philosophie de la systématique ». Le second axe vise à exporter les outils d’analyse historique des structures du vivant vers d’autres disciplines. Par exemple, les linguistes utilisent depuis quelques années les concepts et les outils de la systématique phylogénétique pour inférer l’histoire des langues à partir de leur structure. De même, avec Chomin Cunchillos, j’ai exporté ces méthodes vers la biochimie lorsqu’il s’est agit d’analyser la structure du métabolisme cellulaire en termes d’histoire évolutive ; ou encore avec Erick Denamur adapté ces méthodes à des organismes produisant de nombreux transferts horizontaux, comme par exemple la phylogénie au sein de l’espèce bactérienne Eschericha coli. Enfin, avec Marie Fisler nous nous sommes engagés sur une recherche de catégorisation des idées des auteurs naturalistes au sujet de "l'arbre du vivant" entre 1766 et 1991 qui utilise les outils de maximisation de cohérence par parcimonie dans le partage des idées, recherche qui conduit un "arbre des arbres".
Des organismes Sur un terrain plus zoologique, mes chantiers sont au nombre de deux : les téléostéens acanthomorphes (17 000 espèces, « poissons » modernes à nageoires épineuses) et les notothénioïdes (134 espèces inféodées à l’océan austral, et qui appartiennent aux acanthomorphes).
Concernant les
notothénioïdes, notre équipe est reconnue internationalement sur la
phylogénie du groupe, sur laquelle nous avons régulièrement publié
depuis 15 ans et encadré quatre DEA/M2 (Wei-Jen Chen (1997), Nicolas
Derome (1999), Sophie Sanchez (2003), Mohamed Berkani (2008)).
Des perspectives Mes perspectives scientifiques concernent le redéploiement de l’interaction anatomie comparée – phylogénie moléculaire au service de cet immense chantier phylogénétique qu’est la phylogénie des acanthomorphes. Mais bien au-delà des seuls téléostéens, il faut que l’anatomie comparée renaisse dans l’établissement où elle a vu le jour et qui est le seul à pouvoir la maintenir. L’avenir des musées d’histoire naturelle ne saurait résider dans la seule possession de moyens de séquençage et d’analyse phylogénétique de séquences, mais dans la pleine interaction scientifique de ceux-ci avec l'anatomie, la morphologie des espèces, et avec les collections d'histoire naturelle.
Dernière actualisation : Août 2013 Réalisation du site : Labsynth.com |